8
The effect of plant density on light interception
in hemp
(Cannabis sativa L.)
Hayo M.G. van der Werf 1*
1 DLO Research Institute for
Agrobiology and Soil Fertility (AB-DLO), Wageningen, The Netherlands
* Present address: INRA,
Laboratoire d’Agronomie, 28 rue de Herrlisheim, BP 507, 68021 Colmar, France.
email: vdwerf@colmar.inra.fr
van der Werf, Hayo M. G. 1997. The
effect of plant density on light interception in hemp (Cannabis sativa L.) Journal
of the International Hemp Association 4(1): 8-13.
The effect of plant density on canopy light
interception and on flowering was investigated in a hemp (Cannabis sativa L.) cv.
‘Kompolti Hibrid TC’ crop grown at initial densities of 10, 30, 90 and 270
plants/m2 in field experiments in 1991 and 1992 in the
Netherlands. Rate of canopy establishment increased with increasing plant
density in both years. In 1991, interception of 90% of light was attained at 537
degree days (base 2°C) from emergence for the crop grown at 10 plants/m2
and at 265 degree days for the crop grown at 270 plants/m2.
In 1992, rate of canopy establishment was slower, in particular at 90 and 270
plants/m2, probably as a result of water stress. Maximum
light interception did not depend on plant density and was about 99%. In both years,
the flowering date was later with increasing plant density. Dates of 75% flowering
for the initial densities of 10, 30, 90 and 270 plants/m2
were respectively 11, 17, 20 and 24 August. Independent of plant density, canopy
light interception started to decline at about 400 degree days after flowering, reaching
70% at about 700 degree days post-flowering. The results presented here can be used
to account for the effect of a wide range of plant densities in the simulation of the
course of light interception by a hemp crop.
Introduction
When crops are healthy and
water and nutrients are not limiting, dry matter production is proportional to the amount
of light intercepted by a crop canopy (Monteith 1977). In North-west Europe,
incoming radiation is greatest in May and June, but the interception of solar radiation by
spring-sown crops is never total, because the canopy is still being established.
Rapid canopy establishment is important, as it contributes to increased total light
interception over the growing season and to effective weed suppression by the crop (Lotz et
al. 1991).
Development and growth of plants and crops can
be related to temperature using thermal time, which is the accumulated temperature above a
base temperature, the latter being the temperature below which the process being studied
apparently stops. In field crops, the course of proportional light interception by
the canopy from plant emergence to full ground cover can be described as a logistic
function of thermal time (Spitters 1990). Previous studies (Meijer et al.
1995, van der Werf 1995a) have shown that this approach can be effective in describing
canopy establishment in hemp. However, these results need to be verified and the
effect of plant density needs to be determined.
At some point after flowering, the fraction of
light intercepted by the hemp canopy starts declining as a result of senescence (van der
Werf et al. 1995b). This “onset of senescence” development
stage has not been quantified for hemp, and it is unknown whether plant density affects
onset of senescence or the rate at which light interception subsequently decreases.
Cannabis crops are grown at a wide
range of plant densities, depending on the goal of production and the expected yield
level. For the production of cannabinoids, a density of 10 plants/m2
may be economically optimal (Rosenthal 1987), for seed production a density of 30 plants/m2 may be used (Hennink et al. 1994, van der Werf
1994), whereas for stem or fibre production, densities between 50 and 750 plants/m2 have been recommended (Dempsey 1975). In order to
obtain data required for a simulation model of hemp growth and yield, we examined the
effect of plant density on the course of light interception by the canopy.
Materials and methods
Field experiments were carried
out at Randwijk, the Netherlands (51° 56’ N, 5° 43’ E; 7 m elevation) on a
heavy clay soil in 1991 and 1992. Meteorological data were measured at the
Wageningen Haarweg meteorological station at 5 km from the experimental site. Daily
mean air temperatures were calculated as the average of daily minimum and maximum air
temperatures. Daily thermal time (t) was calculated as:
t = {(Ta - Tb); or 0 if (Ta < Tb)}
where Ta is the daily mean air temperature and Tb is the base temperature.
We used a base temperature of 0°C for the period between sowing and emergence and
a base temperature of 2°C for the course of canopy light interception, as previous
results (van der Werf et al. 1995a) have shown these base temperatures to be
most appropriate.
The Hungarian cultivar ‘Kompolti Hibrid
TC’ was grown at 10, 30, 90 and 270 plants/m2 in
1991 and at 10, 30, 90 and 230 plants/m2 in 1992.
These treatments will be referred to as ‘TC10’, ‘TC30’,
‘TC90’ and ‘TC270’, respectively. The experiment was set up as a
randomized block design with four replications. In 1992, one replication was
discarded because poor soil structure resulted in severe drought stress. Phosphorus
and potassium fertilizer were applied according to recommendations based on soil testing.
Soil nitrogen between 0 and 60 cm was 70 kg/ha in February 1991 and 90 kg/ha in
March 1992. In both years, nitrogen fertilizer was applied before or shortly after
sowing, at a rate of 140 kg/ha in 1991 and 120 kg/ha in 1992. The seed was treated
with a bird repellent (methiocarb, 5 g/kg) and a fungicide (thiram, 3 g/kg).
Hemp was sown with a precision seed drill on 20
April 1991 and on 24 April 1992. Row width was 25 cm for the treatments TC10 and
TC30, and 12.5 cm for the other treatments. The date of 50% plant emergence was 1
May in 1991 and 2 May in 1992. In the seedling stage, the crops were hand-thinned to
the desired plant density. No herbicides were used, because in most treatments,
weeds were suppressed by the crop. In treatments TC10 and TC30, weeds were removed
by hand when necessary. The crops were sprayed with fungicides (alternating
vinchlozolin and iprodion, both at 500 g/ha) at 14-day intervals during June, July and
August. This prevented the occurrence of Botrytis cinerea in all treatments
but TC10. The plots were 14 m long by 6 m wide, and each plot contained four 1.5 x 1
m sample areas, 2 m apart. Results from destructive samplings have been reported in
van der Werf et al. (1995b). The experiment was surrounded by a 3
m-wide buffer of hemp.
The interception of photosynthetically active
radiation (PAR) by the canopy was measured at intervals of one or two weeks from May 21 to
October 6. We used a line sensor (TFDL-DLO, Wageningen, The Netherlands) and
approximately five measurements were taken per plot. Starting in August, the tops of
10 plants per plot were classified weekly as female (bearing one or more female flowers
with visible styles), male (bearing one or more pedicellate male floral clusters) or
non-flowering. Flowering date was defined as the day at which 75% of plants were
classified as female or male, as estimated by interpolation. We calculated the date
of 75% flowering rather than the date of 50% flowering because data to calculate 50%
flowering were not available for all treatments. In these experiments, 75% flowering
occurred 6 to 9 days (corresponding to 100 to 150 degree days (°Cd), base 2°C) after 50%
flowering.
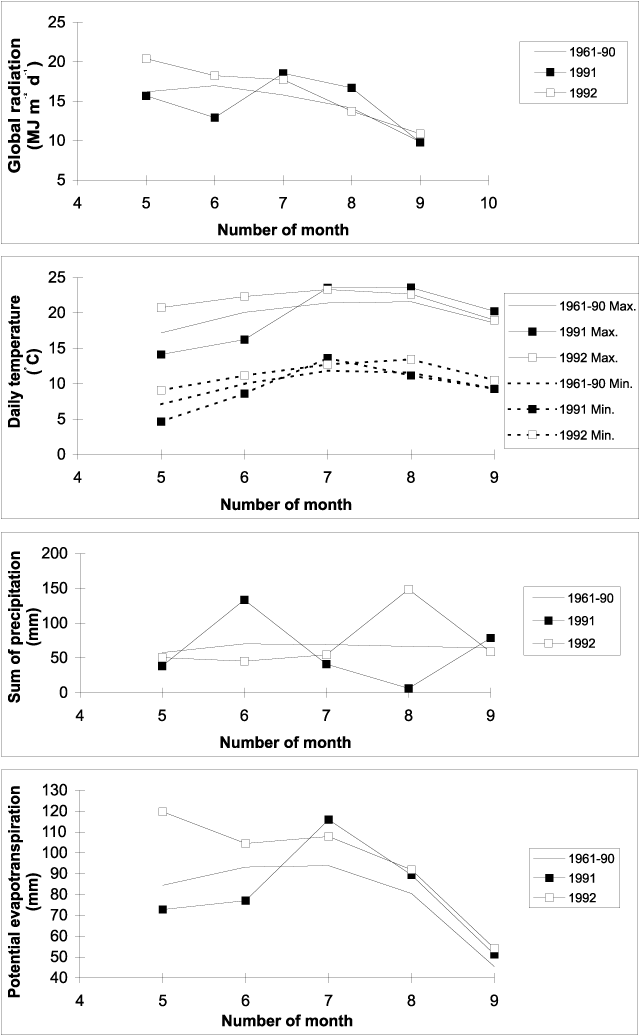
Figure 1. The course of global radiation, daily maximum and minimum air temperature, sum of precipitation and potential evaporation from May to September. Monthly values for 1991, 1992 and the 1961-1990 average, measured at the Wageningen Haarweg meteorological station.
Results and discussion
Weather conditions differed
markedly between the two growing seasons (Fig. 1). Especially for May and June,
which is the period of canopy establishment, global radiation, daily maximum and minimum
temperatures and potential evapo-transpiration were below the benchmark long-term average
in 1991 and well above it in 1992. In May 1992, the evaporative demand was
particularly large.
Canopy establishment
Thermal time (base 0°C)
between sowing and emergence was 62.1°Cd in 1991 and 81.0°Cd in 1992. This is in
reasonable agreement with results obtained in sowing date experiments carried out on a
peaty sand soil where thermal time between sowing and emergence ranged from 68°Cd to
109°Cd, with an average of 88.3°Cd (van der Werf et al. 1995a).
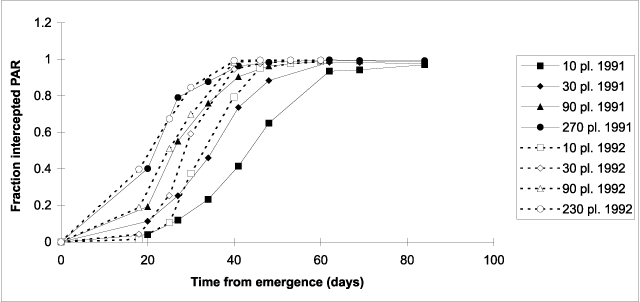
Figure 2. Fraction of photosynthetically active radiation (PAR) intercepted by the canopy of hemp crops versus days from emergence during canopy establishment.
In both 1991 and 1992, the number of days required to reach 90% interception of PAR decreased with increasing plant density (Fig. 2). It ranged from 36 to 60 in 1991 and from 34 to 44 in 1992. For the treatments TC90 and TC270, the time course of light interception was similar in 1991 and 1992. However, for TC10 and TC30, canopy establishment took fewer days in 1992 than in 1991. Maximum light interception was about 99% in both years and did not depend on plant density (data not shown).
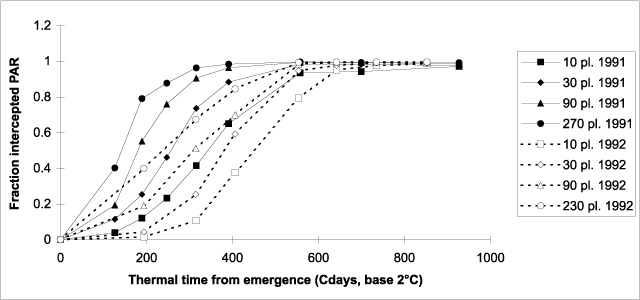
Figure 3. Fraction of photosynthetically active radiation (PAR) intercepted by the canopy of hemp crops versus thermal time (base 2°C) from emergence.
In sowing date experiments carried out during three years, the use of thermal time allowed the accounting of nearly all of the variance in the time course of PAR interception caused by different years and sowing dates (van der Werf et al. 1995a). However, for the experiments reported here, the expression of canopy establishment as a function of thermal time from emergence revealed a pronounced difference: canopy establishment was slower in 1992 than in 1991 for all densities (Fig. 3). Thermal time (base 2°C) required to reach 90% interception of PAR ranged from 265 to 537°Cd in 1991 and from 460 to 613°Cd in 1992. In order to compare the 1991 and 1992 results to those obtained in the sowing date experiments (van der Werf et al. 1995a) and in earlier plant density experiments (Meijer et al. 1995), thermal time from emergence to 90% interception of PAR by the canopy was expressed as a function of plant density (Fig. 4). For this comparison, a base temperature of 0°C was used, as the results of the sowing date experiments and the earlier plant density experiments were available as a function of thermal time with a base of 0°C. Previous results (except for those obtained in 1989) are more in line with the results obtained in 1991 than 1992. In the latter year, rate of canopy establishment expressed as a function of thermal time was slower, in particular for TC90 and TC270 (Fig. 4). This possibly was the result of the high evaporative demand in May 1992 (Fig. 1), which may have caused water stress, slowing down leaf area expansion in the high-density treatments where demand for water was larger.
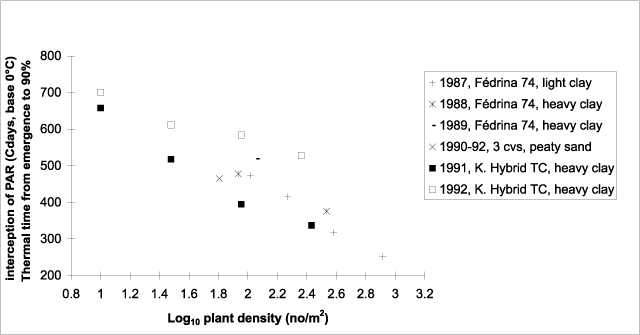
Figure 4. The relationship between thermal time (base 0°C) required from emergence to 90% interception of photosynthetically active radiation (PAR) and log10 of plant density of hemp crops.
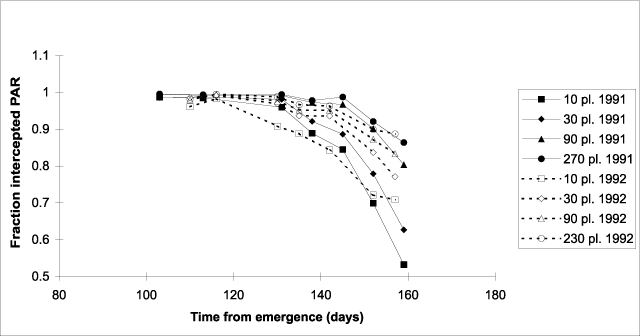
Figure 5. Fraction of photosynthetically active radiation (PAR) intercepted by the canopy of hemp crops versus days from emergence during canopy senescence.
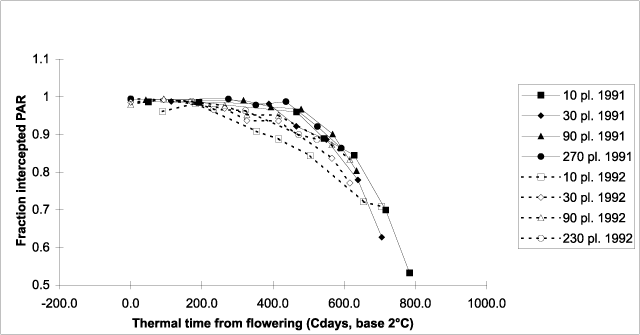
Figure 6. Fraction of photosynthetically active radiation (PAR) intercepted by the canopy of hemp crops versus thermal time (base 2°C) from flowering.
Flowering and senescence
In 1991 and in 1992, flowering
date was later with increasing plant density. For TC10, TC30, TC90 and TC270, dates
of 75% flowering were respectively 9, 14, 19 and 22 August in 1991 and 14, 19, 20 and 26
August in 1992.
A plot of canopy light interception
post-flowering as a function of time from emergence reveals that in both years the onset
of decline of light interception depends in a similar way on plant density: the higher the
plant density, the later light interception starts declining (Fig. 5). When canopy
light interception is expressed as a function of thermal time from 75% flowering,
differences due to plant density disappear, except for TC10 in 1992 (Fig. 6). So the
effect of plant density on the date of onset of senescence apparently resulted mainly from
differences in flowering date associated with differences in plant density. At
400°Cd after flowering, light interception of all treatments (except for TC10 in 1992) is
still approximately 95% or more, afterwards it starts declining, reaching 70% at
approximately 700°Cd post-flowering (Fig. 6).
Conclusions
These experiments have shown
major effects for plant density on rate of canopy establishment and flowering date.
The timing of the onset of senescence does not depend on plant density, but depends on
flowering date only. Provided flowering date is known or can be predicted, these
data can be used to account for the effect of a wide range of plant densities in the
simulation of the course of light interception by a hemp crop.
Acknowledgements
The author thanks M.
Wijlhuizen, J.A.A. de Schutter and H. van der Hoek for technical support in the field and
data analysis.
References
Dempsey J M, 1975. Hemp. Pages 46-89 in Fiber Crops. University of Florida Press, Gainesville, Florida, USA.
Hennink S, de Meijer E P M, van der Werf H M G, 1994. Fiber hemp in the Ukraine, 1991. Pages 261-278 in Rosenthal E (ed.) Hemp Today. Quick American Archives, San Francisco, USA.
Lotz L A, Groeneveld R M W , Habekotte B, van Oene H, 1991. Reduction of growth and reproduction of Cyperus esculentus by specific crops. Weed Research 31: 153-160.
Meijer W J M, van der Werf H M G, Mathijssen E W J M, van den Brink P W M, 1995. Constraints to dry matter production in fibre hemp (Cannabis sativa L.). European Journal of Agronomy 4: 109-117.
Monteith J L, 1977. Climate and the efficiency of crop production in Britain. Philosophical Transactions of the Royal Society of London, B, 281:277-294.
Rosenthal E, 1987. Marijuana Growers Handbook. Indoor/greenhouse edition. Quick American Publishing Company, San Francisco, USA.
Spitters C J T, 1990. Crop growth models, their usefulness and limitations. Acta Horticulturae 267: 349-368.
Van der Werf H M G, 1994. Fiber hemp in France. Pages 213-220 in: Rosenthal E (ed.) Hemp Today. Quick American Archives, San Francisco, USA.
Van der Werf H M G, Brouwer K, Wijlhuizen M, Withagen J C M, 1995a. The effect of temperature on leaf appearance and canopy establishment in fibre hemp (Cannabis sativa L.). Annals of Applied Biology 126: 551-561.
Van der Werf H M G, Wijlhuizen M, de Schutter J A A, 1995b. Plant density and self-thinning affect yield and quality of fibre hemp (Cannabis sativa L.). Field Crops Research 40: 153-164.
Van der Werf H M G, Mathijssen E W J M, Haverkort A J, 1996. The potential of hemp (Cannabis sativa L.) for sustainable fibre production: a crop physiological appraisal. Annals of Applied Biology 129: 109-123.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}